C15:0 (Fatty15, Pentadecanoic acid) a fatty acid from grass-fed animals may be essential - several studies
C-15:0 may be an essential fatty acid - review Dec 2024
New insights on pentadecanoic acid with special focus on its controversial essentiality: A mini-review
Biochimie Volume 227, Part B, Dec 2024, Pages 123-129 https://doi.org/10.1016/j.biochi.2024.10.008
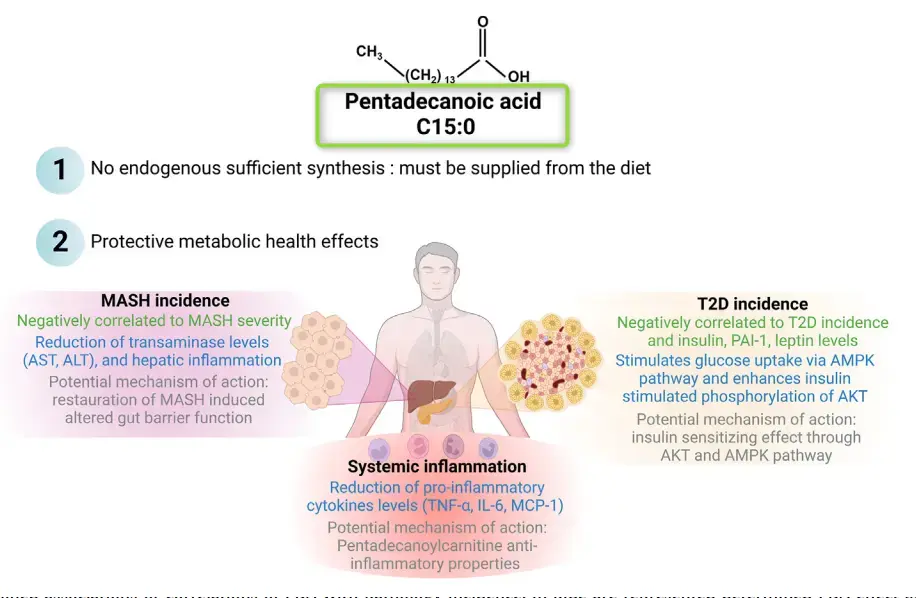
Pentadecanoic acid (C15:0, PDA ) is an odd and minor fatty acid that has been neglected in the literature until the last decade. Indeed, as a specific fatty acid of dairy fat, PDA was only used as a biomarker of dairy fat consumption. Lately, PDA was first correlated negatively with the incidence of metabolic syndrome disorder, then its physiological effects have been investigated as a protective fatty acid. PDA supplementation has been demonstrated as negatively correlated with elevated levels of leptin, plasminogen activator inhibitor-1 and insulin, and has been shown to exhibit sensitizing insulin effects with activation of AMPK pathway. PDA also reduced the severity of metabolic dysfunction-associated steatohepatitis (MASH), notably through reduced alanine transaminase and pro-inflammatory cytokines levels. The final effect described for PDA is its ability to display anti-inflammatory properties in several pathology models. Hence, considering these multiple effects, the presence of PDA could be associated with a healthier physiological state, this raises the question of whether the presence of PDA in the body, in adequate quantities, is needed to participate to health maintenance. PDA is not synthesized in sufficient quantities endogenously, so it must be provided by the diet, mainly through dairy fat, although other types of food can also contribute to the dietary intake of PDA . Essential fatty acids are described as not being endogenously synthesized in sufficient and required quantities to maintain physiological health.
Thus, PDA might gather both conditions to be described as essential, yet further investigations on both criteria are needed to enhance knowledge on this odd chain fatty acid with promising impact as potential protective supplement nutrient.
📄 Download the PDF from VitaminDWiki
C15:0 — Found in Dairy — May Be an Essential Fat - Mercola Aug 2024
Includes a 1 hour video by Venn-Watson
" C15:0, also known as pentadecanoic acid , is an odd-chain saturated fat primarily found in dairy products, some fish, and certain plants. The story of C15:0's importance begins in an unlikely place — with dolphins."
"Dr. Stephanie Venn-Watson , a veterinary epidemiologist, was brought on board the Navy's Marine Mammal program about 20 years ago to help understand aging in dolphins and protect their health. The Navy has been caring for a population of dolphins for over 60 years, and these dolphins are living much longer in captivity (40 to 50+ years) compared to their wild counterparts (around 20 years)."
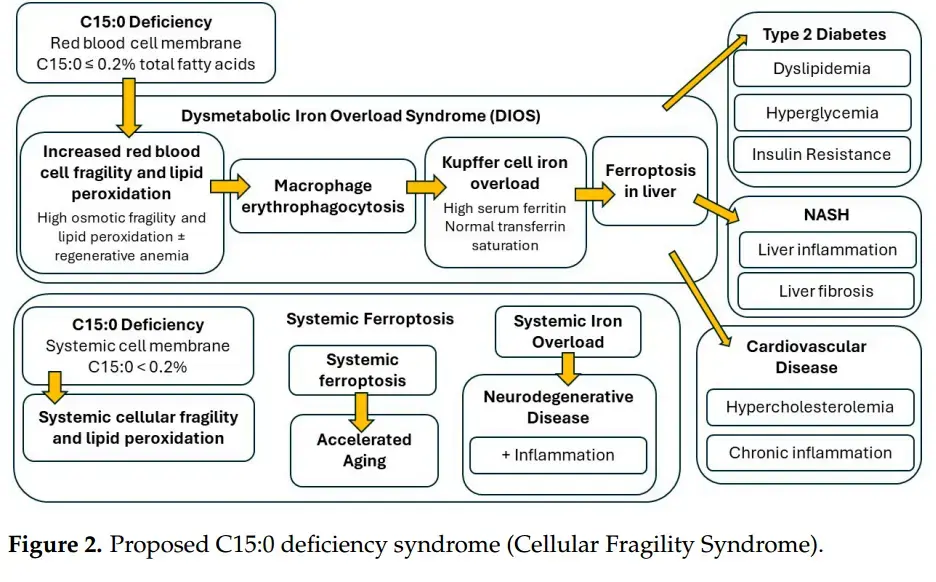
"Dr. Venn-Watson's research suggests that when your C15:0 levels drop below 0.2% of total fatty acids in your cell membranes, you enter a state she calls "Cellular Fragility Syndrome." This syndrome is characterized by fragile red blood cells, anemia, iron overload in the liver, and increased risk of conditions like Type 2 diabetes, cardiovascular disease and fatty liver disease. It's a domino effect that starts at the cellular level and cascades into systemic health issues."
"The impact of C15:0 deficiency is far-reaching and multifaceted. The "Cellular Fragility Syndrome" resulting from C15:0 deficiency is characterized by a cascade of health issues. It starts with fragile red blood cells susceptible to lipid peroxidation, leading to anemia and dysmetabolic iron overload syndrome (DIOS). This iron overload can trigger ferroptosis in the liver, potentially leading to advanced nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH)."
"The syndrome also encompasses insulin resistance, metabolic syndrome, Type 2 diabetes, and cardiovascular disease. Perhaps most alarmingly, it includes systemic iron overload and ferroptosis, which can accelerate aging and tissue damage throughout your body. It's a complex web of interconnected health issues, all potentially stemming from a deficiency in this one crucial fat."
"Based on the available research, your cell membranes require more than 0.2% C15:0 to ensure cellular stability. Optimal circulating C15:0 concentrations should be between 0.4% to 0.64% of total fatty acids. C15:0 deficiency is defined as 0.21% or less of total circulating fatty acids"

The Cellular Stability Hypothesis: Evidence of Ferroptosis and Accelerated Aging-Associated Diseases as Newly Identified Nutritional Pentadecanoic Acid (C15:0) Deficiency Syndrome - June 2024
Metabolites 2024, 14(7), 355; https://doi.org/10.3390/metabo14070355
by Stephanie Venn-Watson 1,2ORCID
1 Seraphina Therapeutics Inc., San Diego, CA 92106, USA
2 Epitracker Inc., San Diego, CA 92106, USA
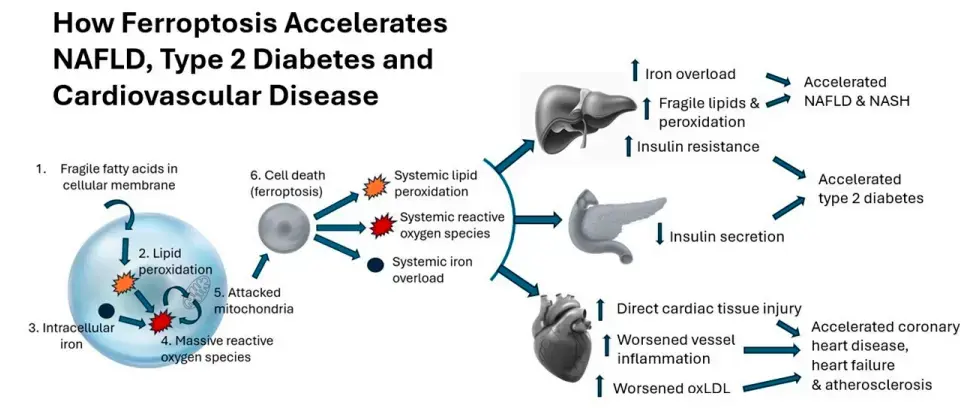
Ferroptosis is a newly discovered form of cell death caused by the peroxidation of fragile fatty acids in cell membranes, which combines with iron to increase reactive oxygen species and disable mitochondria. Ferroptosis has been linked to aging-related conditions, including type 2 diabetes, cardiovascular disease, and nonalcoholic fatty liver disease (NAFLD). Pentadecanoic acid (C15:0), an odd-chain saturated fat, is an essential fatty acid with the primary roles of stabilizing cell membranes and repairing mitochondrial function. By doing so, C15:0 reverses the underpinnings of ferroptosis.
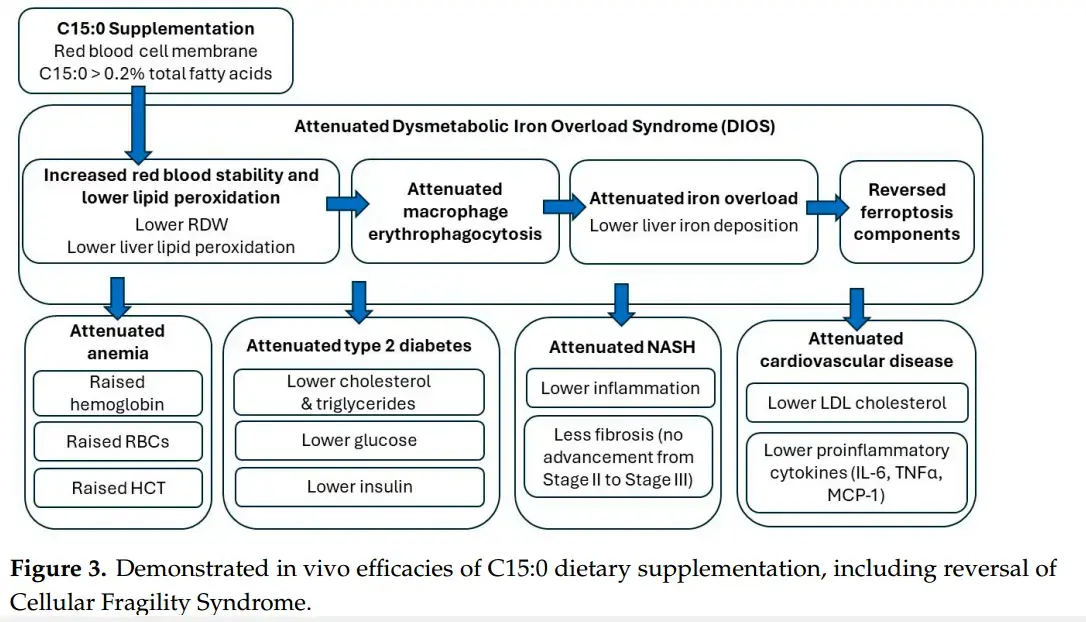
Under the proposed “Cellular Stability Hypothesis”, evidence is provided to show that cell membranes optimally need >0.4% to 0.64% C15:0 to support long-term health and longevity. A pathophysiology of a newly identified nutritional C15:0 deficiency syndrome (“Cellular Fragility Syndrome”) is provided that demonstrates how C15:0 deficiencies (≤0.2% total circulating fatty acids) can increase susceptibilities to ferroptosis, dysmetabolic iron overload syndrome, type 2 diabetes, cardiovascular disease, and NAFLD. Further, evidence is provided that C15:0 supplementation can reverse the described C15:0 deficiency syndrome, including the key components of ferroptosis. Given the declining dietary intake of C15:0, especially among younger generations, there is a need for extensive studies to understand the potential breadth of Cellular Fragility Syndrome across populations.
📄 Download the PDF from VitaminDWiki
Mercola webpage - June 2026
There are several of vendors on Amazon for those who do not get grass-fed dairy or meat (goats, cows, etc.)
Note: Cost/capsule varies by 20X: $0.08 to $1.67
Broader and safer clinically-relevant activities of pentadecanoic acid compared to omega-3: Evaluation of an emerging essential fatty acid across twelve primary human cell-based disease systems - May 2022
Stephanie K. Venn-Watson , Camden N. Butterworth
PLOS x https://doi.org/10.1371/journal.pone.0268778
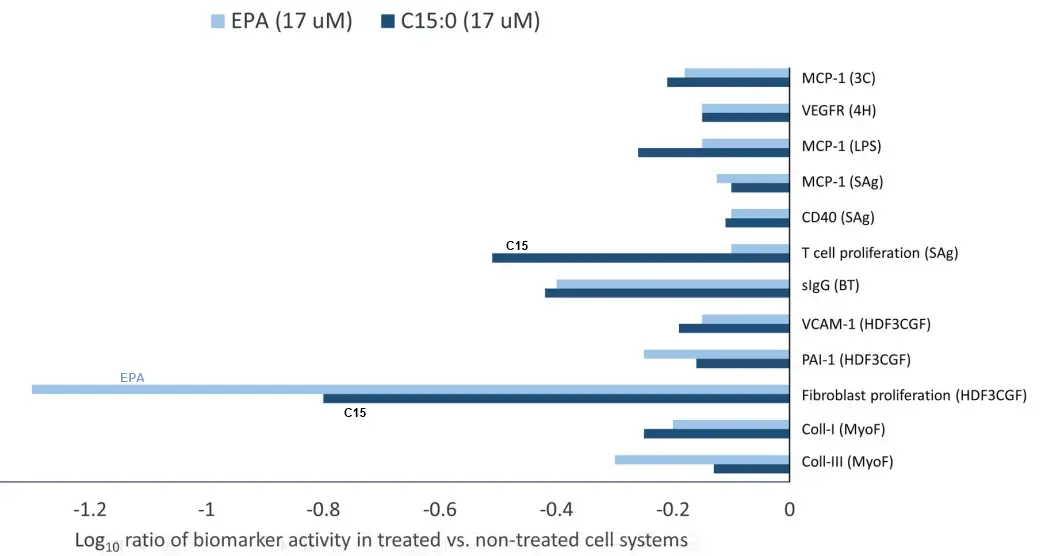
A growing body of evidence supports that pentadecanoic acid (C15:0), an odd-chain saturated fat found in butter, is an essential fatty acid that is necessary in the diet to support long-term metabolic and heart health. Here, dose dependent and clinically relevant cell-based activities of pure C15:0 (FA15TM) were compared to eicosapentaenoic acid (EPA), a leading omega-3 fatty acid, as well as to an additional 4,500 compounds. These studies included 148 clinically relevant biomarkers measured across 12 primary human cell systems, mimicking various disease states, that were treated with C15:0 at four different concentrations (1.9 to 50 μM) and compared to non-treated control systems. C15:0 was non-cytotoxic at all concentrations and had dose dependent, broad anti-inflammatory and antiproliferative activities involving 36 biomarkers across 10 systems. In contrast, EPA was cytotoxic to four cell systems at 50 μM. While 12 clinically relevant activities were shared between C15:0 and EPA at 17 μM, C15:0 had an additional 28 clinically relevant activities, especially anti-inflammatory, that were not present in EPA. Further, at 1.9 and 5.6 μM, C15:0 had cell-based properties similar to bupropion (Pearson’s scores of 0.78), a compound commonly used to treat depression and other mood disorders. At 5.6 μM, C15:0 mimicked two antimicrobials, climabazole and clarithromycin (Pearson’s scores of 0.76 and 0.75, respectively), and at 50 μM, C15:0 activities matched that of two common anti-cancer therapeutics, gemcitabine and paclitaxel (Pearson’s scores of 0.77 and 0.74, respectively). In summary, C15:0 had dose-dependent and clinically relevant activities across numerous human cell-based systems that were broader and safer than EPA, and C15:0 activities paralleled common therapeutics for mood disorders, microbial infections, and cancer. These studies further support the emerging role of C15:0 as an essential fatty acid.
📄 Download the PDF from VitaminDWiki
Pentadecanoic acid and T2 Diabetes - Dec 2014
Serum pentadecanoic acid (15:0), a short-term marker of dairy food intake, is inversely associated with incident type 2 diabetes and its underlying disorders
The American Journal of Clinical Nutrition Volume 100, Issue 6 , Dec 2014, Pages 1532-1540 https://doi.org/10.3945/ajcn.114.092544
Background:
Growing evidence suggests that dairy consumption is associated with lower type 2 diabetes risk. However, observational studies have reported inconsistent results, and few have examined dairy’s association with the underlying disorders of insulin resistance and β-cell dysfunction.
Objective:
We investigated the association of the dairy fatty acid biomarkers pentadecanoic acid (15:0) and trans-palmitoleic acid (trans 16:1n−7) with type 2 diabetes traits by evaluating 1) prospective associations with incident diabetes after 5 y of follow-up and 2) cross-sectional associations with directly measured insulin resistance and β-cell dysfunction.
Design:
The study analyzed 659 adults without diabetes at baseline from the triethnic multicenter Insulin Resistance Atherosclerosis Study (IRAS). Diabetes status was assessed by using oral-glucose-tolerance tests. Frequently sampled intravenous-glucose-tolerance tests measured insulin sensitivity (SI) and β-cell function [disposition index (DI)]. Serum fatty acids were quantified by using gas chromatography. Logistic and linear regression models were adjusted for demographic, lifestyle, and dietary variables.
Results:
Serum 15:0 was a significant biomarker for total dairy intake in the IRAS cohort. It was associated with a decreased incident diabetes risk (OR: 0.73, P = 0.02) and was positively associated with log SI (β: 0.84, P = 0.03) and log DI (β: 2.21, P = 0.02) in fully adjusted models. trans 16:1n−7 was a marker of total partially hydrogenated dietary fat intake and was not associated with outcomes in fully adjusted models.
Conclusions:
Serum 15:0, a marker of short-term intake of this fatty acid, was inversely associated with diabetes risk in this multiethnic cohort. This study may contribute to future recommendations regarding the benefits of dairy products on type 2 diabetes risk.
📄 Download the PDF from VitaminDWiki
Health benefits of Pentadecanoic acid - Perplexity AI Aug 2024
Pentadecanoic acid (C15:0) is an odd-chain saturated fatty acid that has been recognized for its potential health benefits. Here are some of the key health benefits associated with pentadecanoic acid:
Cardiometabolic Health : C15:0 is linked to improved cardiometabolic health, including lower risks of type 2 diabetes, heart disease, and nonalcoholic fatty liver disease. It helps in reducing cholesterol and triglyceride levels, which are crucial for cardiovascular health.
Anti-inflammatory and Antifibrotic Effects : C15:0 exhibits anti-inflammatory and antifibrotic properties. It activates AMPK and PPAR-α/δ, which are involved in metabolic regulation, and inhibits mTOR, JAK-STAT, and HDAC-6 pathways, contributing to its anti-inflammatory effects.
Longevity and Aging : Studies suggest that C15:0 may enhance healthspan and longevity. It has been compared to rapamycin, a known longevity-enhancing drug, for its cell-repairing benefits and potential to delay the onset of age-related diseases.
Metabolic Regulation : C15:0 supports metabolic health by improving mitochondrial function, stabilizing red blood cells, and regulating glucose metabolism. These activities help maintain healthy glucose and cholesterol levels.
Cellular Health : C15:0 enhances cellular resilience by incorporating into cell membranes, improving their stability and function. It also helps in maintaining cellular communication by activating PPARs, which are crucial for metabolism and immunity.
Potential Antimicrobial and Antidepressant Properties : C15:0 has shown antimicrobial effects against certain bacteria and fungi, and its properties have been likened to those of antidepressants, suggesting potential benefits for mood disorders.
Overall, pentadecanoic acid is emerging as a nutrient with broad health benefits, particularly in supporting metabolic health, reducing inflammation, and promoting longevity.
C15:0 Review by Claude AI April 2026
C15:0 is an odd-chain saturated fatty acid with 15 carbons. It's found in trace amounts in dairy fat (about 1-3% of butterfat), ruminant meat, and some fish and plants. It's drawn substantial attention since around 2020 because a research group has argued it qualifies as a previously unrecognized essential fatty acid.
The origin story. The hypothesis emerged from veterinary work. Stephanie Venn-Watson, a veterinary epidemiologist, was studying Navy dolphins and noticed that healthier, longer-lived dolphins consumed more C15:0 — not, as expected, more omega-3s. After the rodent and cell work that followed, she co-founded the supplement Fatty15.
The case for C15:0 mattering in humans. Several large prospective cohorts have found that higher blood concentrations of C15:0 track with lower risk of type 2 diabetes, cardiovascular disease, and heart failure, and that higher dietary intake and circulating concentrations correlate with lower mortality, less chronic inflammation, lower rates of gestational diabetes, hypertension, NAFLD, NASH, and COPD. Mechanistically, the published work describes C15:0 as a dual partial PPARα/δ agonist, AMPK activator, and HDAC6 inhibitor that appears to repair mitochondrial function, stabilize red blood cells, regulate glucose metabolism, and reduce cancer cell proliferation. In cell-based phenotyping assays, it shows overlapping activity profiles with rapamycin (mTOR inhibition) and metformin (AMPK activation) — both heavily studied longevity candidates. The proposed dose is 100 to 300 mg/day to maintain circulating concentrations of roughly 10 to 30 µM.
The "essential" argument rests on three legs: humans don't readily synthesize C15:0 endogenously; population C15:0 levels have been declining for decades alongside the shift away from full-fat dairy; and lower levels predict worse outcomes.
The skeptical view, which is worth weighing seriously. No major medical or regulatory body has classified C15:0 as essential. Most of the foundational human research traces back to one group with a direct commercial stake (Seraphina Therapeutics / Fatty15). Population correlations are confounded — C15:0 is a biomarker of dairy fat intake, which correlates with many other dietary and lifestyle variables. The marketed "Cellular Fragility Syndrome" and the "1 in 3 are deficient" framing aren't recognized clinical entities; they're constructs built on the company's own threshold definitions. Bill Harris, a well-regarded omega-3 researcher, has publicly questioned whether the marketing is running ahead of the evidence, particularly the framing of C15:0 as comparable to or superior to omega-3s, where the evidence base is far thinner and the independent RCTs are largely missing.
A frame you might find familiar. This case is in some ways an inversion of the "profitable ignorance" pattern. Here someone is trying to commercialize a cheap, naturally occurring molecule — but in doing so they've patented a purified form, generated most of the existing literature in-house, and built a marketing apparatus around a not-yet-established essentiality claim. The biology may well turn out to be real (the in vitro signal is genuinely interesting), but the gap between "biologically active small molecule" and "officially essential nutrient with deficiency syndromes" is large, and that gap is currently being filled by promotional rather than independent science.
Practical bottom line. If the dairy-fat hypothesis is right, full-fat dairy, butter, and ruminant meat are the cheapest route to meaningful intake — a few hundred mg/day is achievable through diet. The pure supplement form is an option for people avoiding dairy, but at current pricing it's a bet on an essentiality claim that hasn't been independently validated. For someone tracking the mitochondrial / AMPK / mTOR longevity literature, C15:0 is worth keeping an eye on — but the prudent stance is "promising and biologically plausible, not established."
C15:0 development: similar to Vitamin D in 1921 - Claude AI April 2026
Where C15:0 sits on the timeline. The honest comparison is that C15:0 today is somewhere around where vitamin D was in 1920–1922 — Mellanby had shown a fat-soluble factor cured rickets in dogs (1919), McCollum was sorting out vitamin A from this newer factor (1922), but essentiality wasn't yet broadly accepted, the molecule wasn't characterized, and you had a small number of researchers driving the agenda. That's the most generous mapping. A less generous mapping is that C15:0 is where vitamin K2 was about 15 years ago, or where CoQ10 was in the late 1990s — bioactive small molecule, suggestive cohort data, articulate advocates, commercial interest, no recognized deficiency syndrome.
The reason C15:0 doesn't map cleanly onto, say, 1990s vitamin D — even though the marketing borrows that vibe — comes down to several structural differences that matter epistemically.
1. Bedrock of essentiality. Vitamin D had rickets. A dramatic, visible, reversible deficiency disease, established before 1925, with a clear dose-response and a clean mechanism via calcium handling. Everything in the modern vitamin D literature — the cancer hypotheses, the immune work, the all-cause mortality signals — gets to stand on top of that bedrock. C15:0 has no analogous foundation. "Cellular Fragility Syndrome" is a phrase coined by the company selling the supplement, not a recognized clinical entity with a phenotype, a diagnostic test, or independent characterization. That's a categorical difference, not a stage difference.
2. Independence of the research base. By 1980, vitamin D had been worked on by hundreds of independent labs across multiple countries for half a century — Mellanby, McCollum, Windaus (Nobel 1928), DeLuca, Holick, Norman, the Garlands, and so on, with no shared commercial interest. The foundational C15:0 essentiality claim is largely concentrated in Venn-Watson and collaborators, with Seraphina Therapeutics in the background. Even where independent groups have measured C15:0 in cohorts (EPIC, PURE, etc.), they originally measured it as a biomarker of dairy fat intake, not as a candidate essential nutrient — the reinterpretation as evidence for essentiality is post-hoc.
3. Receptor and mechanism maturity. The VDR was cloned in 1987; tissue distribution mapping followed quickly; the genomic and non-genomic action models were independently replicated by multiple groups. C15:0's mechanism — partial PPARα/δ agonism, AMPK activation, HDAC6 inhibition — is plausible and increasingly characterized, but it's at an earlier stage and again much of it traces back to the same network.
4. Biomarker quality. 25(OH)D is a well-validated status biomarker with a stable half-life, established assay standardization (after a long fight), and reproducible associations across populations. Circulating C15:0 is much more strongly determined by recent dairy intake; the "deficiency" thresholds are essentially set by the supplement maker rather than derived from independent consensus.
5. RCT base. Vitamin D in 2020 had thousands of RCTs across dozens of indications, with the disappointments (VITAL, D2d, etc.) at least as instructive as the wins. C15:0 has effectively no rigorous independent RCTs for its proposed indications. Most human-relevant evidence is observational or in vitro.
6. Commercial structure. This is where the comparison genuinely flips. Vitamin D is a commodity — unpatentable, cheap, no moat. The "profitable ignorance" pattern you've used elsewhere fits well there: nobody had a strong commercial incentive to push it, so it took decades for the wider claims to get traction. C15:0 has the opposite problem: a patented purified form, a brand, a story, and substantial incentive to generate and amplify favorable literature ahead of the independent evidence catching up. Both patterns distort science, in opposite directions.
One honest caveat. The extra-skeletal vitamin D claims — cancer prevention, autoimmune protection, all-cause mortality reduction — face many of the same epistemological challenges C15:0 does today: heavy reliance on observational data, plausible mechanisms, RCTs that often underperform expectations. So a fair-minded skeptic of C15:0 should also be honest about which parts of the vitamin D consensus rest on the same kind of evidence they're criticizing C15:0 for. The difference is that vitamin D's core essentiality is unimpeachable, so even if every extra-skeletal claim collapsed tomorrow, you'd still need it. With C15:0, if essentiality doesn't hold, there's no fallback.
Bottom line for your evaluation framework. If you used the same epistemic standards on C15:0 today that the medical establishment used on vitamin D's extra-skeletal claims circa 2010, C15:0 would not yet clear the bar. If you used the same standards that you would use — which I'd characterize as "take the mechanism seriously, weight the cohort data, be patient with the RCT gap, watch for profitable ignorance" — then C15:0 is genuinely interesting but earlier-stage, and the commercial pattern around it is in the opposite direction from the one you usually have to fight.